
I w sumie na tym stwierdzeniu można by poprzestać, ale spróbujmy przyjrzeć się problemowi trochę dokładniej.
Aminy – kilka słów wstępu
Pojęcie „amina” używane jest w chemii dla określenia związków przynależnych do pewnej określonej klasy – do amin właśnie. Aminy spełniają następujące kryteria:
1. zawierają atom azotu o hybrydyzacji sp3 – oznacza to, że geometria atomu azotu jest tetraedryczna, trzy naroża tetraedru zajmują podstawniki (atomy węgla lub wodoru), czwarty to wolna para elektronowa atomu azotu (na rysunkach wolną parę elektronową często się pomija);
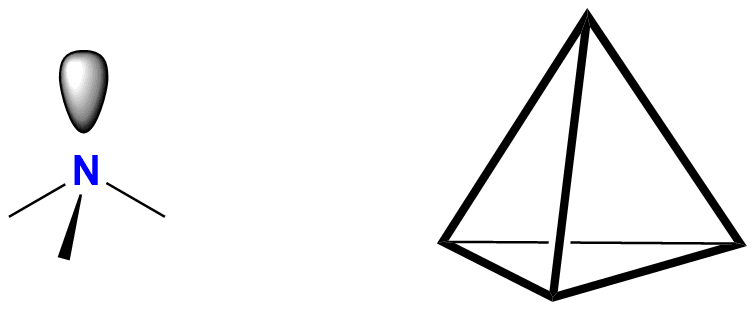
2. atom azotu jest trójwiązalny (jest połączony z trzema różnymi atomami);
3. z atomem azotu związany jest jeden, dwa lub trzy atomy węgla, przy czym muszą to być grupy alkilowe (np.: metylowa, etylowa itd.), alkenowe (np.: winyl), alkinowe (np.: etynylowa) lub arylowe (np.: fenyl).
W zależności od ilości atomów wodoru związanych z atomem azotu mówimy o aminach pierwszorzędowych (dwa atomy wodoru, jeden atom węgla), drugorzędowych (jeden atom wodoru, dwa atomy węgla) i trzeciorzędowych (trzy atomy węgla).
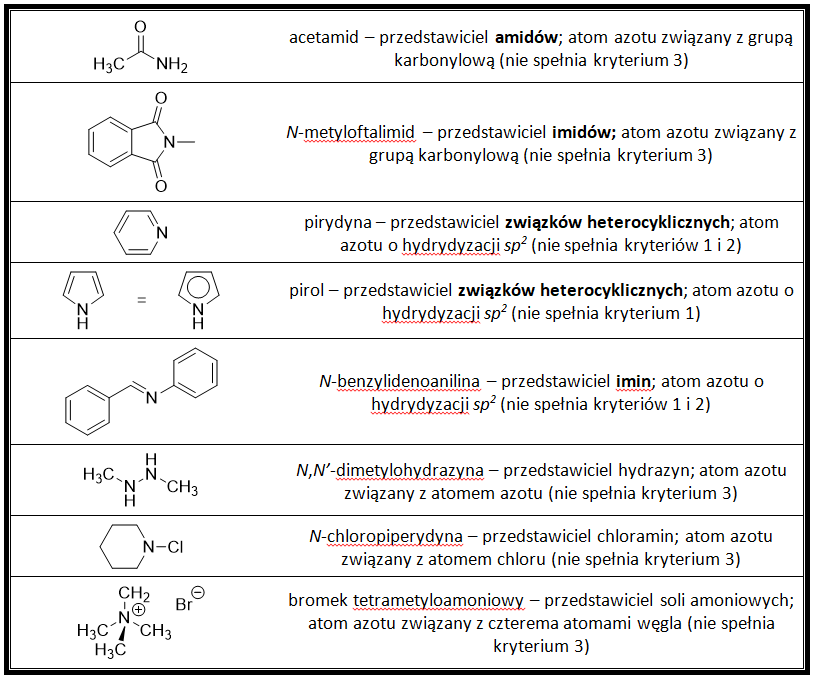
Przykładowe aminy

Przykładowe nie-aminy
Aminy mają charakter zasadowy – mogą przyłączać jon H+ (ulegać protonowaniu). Wiele amin ulega tej reakcji w roztworach wodnych, „zabierając” jon H+ cząsteczce wody (jednocześnie pojawiają się jony hydroksylowe, a roztwór przyjmuje odczyn zasadowy):
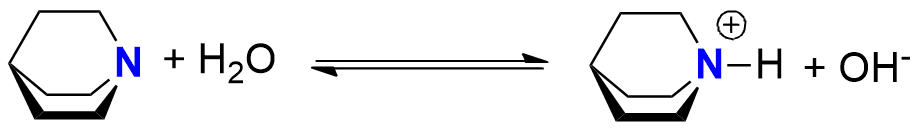
Nie jest to jednak cecha wszystkich amin – niektóre w roztworach wodnych zachowują się jak donor protonu (protonodawca), a ich roztwory mają charakter kwaśny. Takie aminy ulegają protonowaniu tylko w roztworach silnych kwasów.

Aminy mają olbrzymie znaczenie biologiczne – wiele z nich ma kluczowe w funkcjonowaniu ludzkiego organizmu, jest lekami, toksynami, substancjami obronnymi zwierząt i roślin etc. Ale o tym napiszemy przy innej okazji.
Witaminy – trochę historii
Pojęcie „witamina” do języka nauki i życia codziennego wprowadził w 1912 roku Polak pracujący w Londynie (w Instytucie Listera), biochemik, Kazimierz Funk. Ale po kolei…
Do odkrycia witamin doszło podczas badania przyczyn zaburzeń neurologicznych objawiających się polineuropatią obwodową (zaburzenia czuciowe i ruchowe), porażeniami i problemami kardiologicznymi. Choroba ta znana jest pod nazwą beri-beri. W latach 80’ XIX wieku japoński lekarz wojskowy Takaki Kanehiro odrzucił teorię infekcji w etiologii tego schorzenia i powiązał występowanie beri-beri z ubogą dietą. Późniejsze obserwacje (koniec XIX w.) duńskiego patologa, Christiaana Eijkmana dowiodły, że u zwierząt dieta oparta na łuskanym ryżu sprzyja wystąpieniu choroby, natomiast karmienie ryżem niełuskanym jej zapobiega. Pierwotny wniosek z badań był błędny – Eijkman wiązał objawy z toksycznością dużych dawek skrobi, głównego składnika łuskanego ryżu. A nieznany składnik otrąb miał tę toksyczność niwelować. Frederick Hopkins postuluje (1898), że obok cukrów, białek, tłuszczów i składników mineralnych niektóre pokarmy zawierają inne substancje niezbędne do życia (nazywa je „czynnikami dodatkowymi” – „accessory factors”). Rok 1901 przynosi wniosek sformułowany przez Gerrita Grijsa i Ch. Eijkmana – beri-beri to choroba wynikająca z niedoboru jakiegoś składnika pokarmowego. Dziesięć lat później Edward B. Vedder ustala, że ekstrakt z otrąb ryżowych jest lekiem na beri-beri. Prace Hopkinsa i Eijkmana zostały uhonorowane Nagrodą Nobla z fizjologii i medycyny w 1929 roku.
Funk zaczyna badania od ekstrakcji otrąb zakwaszonym etanolem, oddzieleniem frakcji lipidowej i wydzielenia składnika czynnego przez sekwencyjne wytrącanie kwasem fosforowolframowym, chlorkiem rtęci(II) i azotanem(V) srebra.
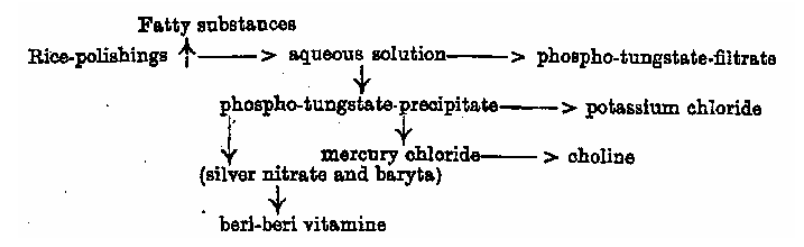
Z 1 kg otrąb uzyskuje 0,5 g substancji wykazującej aktywność przeciw beri-beri w testach na zwierzętach. Funk ustala także wzór sumaryczny badanego związku (błędnie; dziś trwają dyskusje, co w rzeczywistości wyizolował). I stwierdza, opierając się na właściwościach chemicznych, że otrzymany związek jest aminą. Aminą niezbędną do życia, a więc witaminą (łac. „vita” – życie). Cóż, gdyby zaczął od badań nad składnikiem anty-szkorbutowym, to idąc tym tokiem rozumowania mielibyśmy dzisiaj „witacydy”.
Co warte podkreślenia, podobne badania prowadził i opublikował w 1911 r. (a więc przed Funkiem) japoński badacz Umetaro Suzuki. Niestety, jego prace nie przebiły się do międzynarodowego obiegu ze względu na barierę językową.

Struktura witaminy przeciwdziałającej beri-beri została ustalona dopiero w 1936 r. przez Roberta R. Williamsa.
Z powyższej historii wynika kilka ciekawych wniosków, z których dwa najważniejsze można sformułować następująco: (i) odkrycie naukowe nigdy nie jest zawieszone w próżni – wynika z odkryć poprzedników i staje się inspiracją dla następców; (ii) unifikacja języka nauki nie jest taka zła (wbrew utyskiwaniom orędowników „nauki narodowej”) – przynajmniej nie musimy się uczyć japońskiego.
Witaminy – z czym to się je?
Podobno najczęściej z zieleniną. I tłustymi rybami morskimi. Ale spójrzmy na problem oczami chemika. A więc – co robią witaminy na poziomie molekularnym? Pytanie tak postawione nie ma prostej odpowiedzi. Witaminy to grupa związków o różnym charakterze działania:
a. Witaminy stanowiące kofaktory i kosubstraty enzymów
Enzym to po prostu „zwitek białka” – cząsteczka złożona z jednego lub kilku łańcuchów polipeptydowych poskręcanych w przestrzeni. Jej struktura decyduje o właściwościach enzymu. Ale do osiągnięcia najważniejszej własności enzymów, ich aktywności katalitycznej (czyli przyspieszania przekształcania jednych substancji w inne), samo białko w wielu wypadkach nie wystarczy. W takich przypadkach w centrum aktywnym cząsteczki enzymu znajduje się tzw. kofaktor. Jest to jon lub cząsteczka związana z białkiem wiązaniem kowalencyjnym lub słabszymi oddziaływaniami, odpowiadająca za właściwą aktywność enzymu, czyli odwalająca (prawie) całą robotę. Kofaktorem może być jon metalu (np.: Zn2+), kompleks metalu (np.: hem), cząsteczka organiczna. Czasem enzym, aby zadziałać, wymaga kosubstratu – cząsteczki okresowo wiązanej z enzymem, tylko na czas katalizowania reakcji. W roli kofaktorów/kosubstratów często występują witaminy (czasem po drobnej modyfikacji struktury, np.: dodaniu do cząsteczki reszty kwasu fosforowego(V)): B1, B2, B6, B7, B12, C, K, kwas liponowy, kwas foliowy.
b. Prekursory kofaktorów
Niektóre witaminy, aby stać się kofaktorem enzymu, muszą przejść długą drogę, czytaj: ulec szeregowi modyfikacji chemicznych, znacznie rozbudowujących pierwotną cząsteczkę. Taki los czeka witaminę B2 (ulega przekształceniu w dinukleotyd flawinoadeninowy – FAD), kwas pantotenowy (w organizmie występuje w formie koenzymu A lub panteteiny) i niacyna (przekształcana w NAD i NADP, czyli dinukleotyd nikotynoamidoadeninowy i jego fosforan(V)).
c. Chromofor w białkach wrażliwych na światło
Chromofor to fragment cząsteczki absorbujący światło. Witamina A związana z białkami (opsynami) stanowi barwnik wrażliwy na światło i pozwalający na rejestrację bodźców wzrokowych.
d. Substancje sygnałowe i regulatorowe
Cząsteczki witaminy A mogą wiązać się z receptorami obecnymi w jądrach komórkowych, regulując proces ekspresji genów, czyli to kiedy i jak szybko informacja genetyczna w konkretnym genie zostanie „przepisana” na cząsteczkę białka. Receptory jądrowe to białka związane z DNA, które mogą aktywować lub blokować proces transkrypcji informacji genetycznej. Działanie receptora jądrowego zależy od tego, czy ulegnie związaniu z małą cząsteczką-posłańcem, tzw. ligandem (w tej roli występują m.in. witaminy). Receptory jądrowe zależne od witaminy A określane są akronimami RAR i RXR.
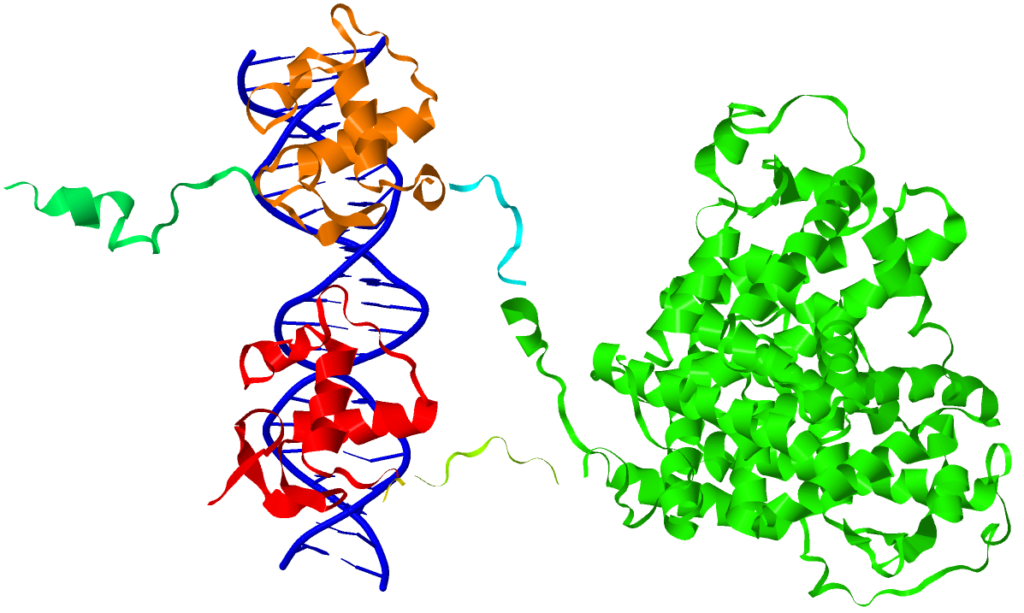
Podobnie działa witamina D – związek ten jest ligandem dla receptora jądrowego VDR. Witamina D wykazuje także inną drogę działania – może wiązać się z receptorem zlokalizowanym w błonie komórkowej, uruchamiając tym samym kaskadę reakcji chemicznych, prowadzących do określonego efektu fizjologicznego (ta droga jest szybsza niż ta, angażująca receptory jądrowe). Tym samym witaminy A i D nie różnią się mechanizmem działania od hormonów.
Witamina E pełni rolę aktywatora enzymu, fosfatazy proteinowej A2, który z kolei wpływa na aktywność innych białek w komórce (poprzez katalizowanie ich defosforylacji, czyli „odcinania” reszt kwasu fosforowego(V)). Dokładny mechanizm oddziaływania witaminy E i fosfatazy nie jest znany (być może znaczenie mają wolne rodniki, których stężenie jest regulowane przez witaminę E).
e. Aktywność antyoksydacyjna i redukcyjna
Witaminy E i C pełnią funkcję „zmiataczy” reaktywnych form tlenu – wolnych rodników i nadtlenków. Nie są podczas tych procesów związane z żadnymi enzymem – działają jako małe, izolowane cząsteczki. Witamina E wbudowuje się (dzięki swojemu powinowactwu do tłuszczów) w podwójne błony lipidowe i chroni te ważne elementy komórki przed uszkodzeniem. Uczestniczy także w ochronie liposomów i lipoprotein. Witamina C odpowiada także za redukcję jonów Fe3+ do Fe2+, na przykład w przewodzie pokarmowym (co ułatwia przyswajanie tego pierwiastka). Literatura wymienia wiele procesów redukcji zachodzących w żywych organizmach pod wpływem witaminy C.
Jak widać z powyższych przykładów nie ma jednego „mechanizmu działania witamin”, a granice między witaminami a innymi klasami związków biologicznie aktywnych są nieostre (stąd zapewne wliczanie do tej grupy przez medycznych szarlatanów różnych dziwnych związków, w przypadku których nie ma przesłanek na ich niezbędność dla organizmu). Należy podkreślić, że powyższe przykłady nie wyczerpują problematyki, a leżące u podstaw „witaminologii” mechanizmy biochemiczne są nadal badane (na przykład w wypadku witaminy B1 istnieją mocne przesłanki na istnienie receptorów dla tego związku, a więc na regulatorowy mechanizm jej działania).
To jak to jest w tymi (wit)aminami?
Tak, zgadliście… odpowiedź ponownie nie jest taka prosta. Wynika to z tego, że każda witamina to w rzeczywistości cała rodzina związków. Dlaczego? Po pierwsze, w organizmie witamina może ulegać przekształceniu w kilka-kilkanaście pochodnych różniących się strukturą. Dlatego jedząc spożywamy różne związki chemiczne z danej grupy, a w dodatku następnie ulegają one przetwarzaniu przez nasze komórki. Przykład? Bardzo proszę – „znana i lubiana” witamina D. W produktach pochodzenia roślinnego występuje „witamina D2”, czyli ergokalcyferol, w produktach odzwierzęcych – „witamina D3”, cholekalcyferol. Co ciekawe, ani jedna, ani druga nie jest właściwą formą aktywną. W tkankach (głównie w wątrobie) obie cząsteczki ulegają hydroksylowaniu (przyłączeniu grupy –OH) w pozycji 25. Powstałe pochodne: 25-hydroksyergokalcyferol i 25-hydroksycholekalcyferol to jeszcze nie właściwe formy czynne. Dopiero po kolejnej hydroksylacji, w pozycji 1, powstają związki o aktywności hormonalnej: 1α,25-dihydroksyergokalcyferol i 1α,25-dihydroksycholekalcyferol. Ten proces zachodzi głównie w nerkach. Jakby tego było mało, w tym samym narządzie może powstawać pochodna 24,25-dihydroksy-, różniąca się nieco aktywnością od izomeru 1α,25-dihydroksy. A gdzie w tym słońce, na którego promienie mamy się wystawiać, aby uniknąć niedoborów witaminy D? U zwierząt kręgowych, a więc i u człowieka, w skórze zachodzi przemiana prowitaminy D3 (czyli 7-dehydrocholesterolu) w prewitaminę D3, a tej w cholekalcyferol (a przy okazji mamy dwie kolejne cząsteczki z rodziny „witamin D”, tym razem z przedrostkami pre- i pro-). W grzybach i roślinach występuje prowitamina D2 (ergosterol), i odpowiadająca jej prewitamina. Mamy jeszcze witaminy D4 i D5. Pierwsza występuje w grzybach, druga w roślinach. No i należy doliczyć całą gamę izomerów, produktów ubocznych, metabolitów będących efektem katabolizmu etc. Duża rodzinka, nieprawdaż?.
„Słowo na dziś”: witamery – formy tej samej witaminy, wykazujące różnice w strukturze cząsteczki, ale cechujące się taką samą lub zbliżoną aktywnością biologiczną lub ulegające przemianie w aktywną formę witaminy.

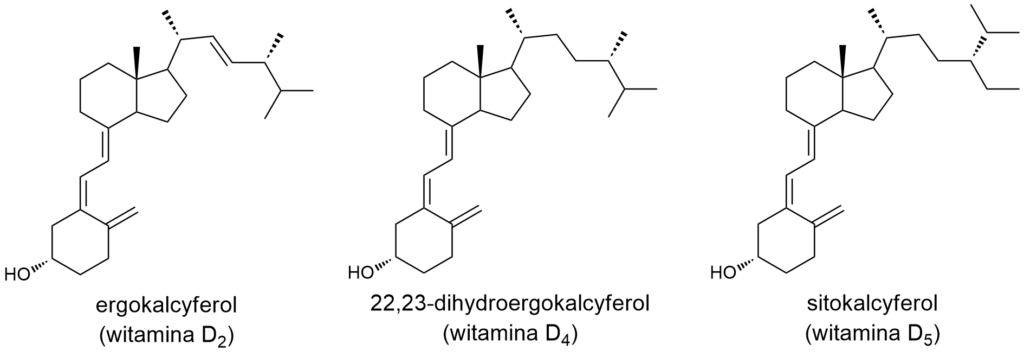
Jak widać na powyższych rysunkach, żadna z witamin D nie jest aminą.
Drugi powód bogactwa związków z grupy witamin to efekt działań przemysłu farmaceutycznego. Produktem stosowanym w lekach, suplementach czy żywności nie musi być cząsteczka tożsama z naturalną witaminą. Często stosuje się pochodną lub analog o takiej samej aktywności, ale łatwiejszy w produkcji, trwalszy, łatwiej wchłanialny etc. Albo poszukuje się związków o wzmocnionej jednej ścieżce działania, a wygaszonej innej.
Doskonałym przykładem takiego podejścia jest witamina B12. W tkankach występuje w postaci metylokobalaminy lub 5’-deoksyadenozylokobalaminy oraz śladowych ilości hydroksykobalaminy, będącej produktem rozkładu dwóch pierwszych związków. Ale główną formą produkowaną i stosowaną w farmacji jest cyjanokobalamina. Dlaczego? Bo jest łatwa do wyizolowania z mieszaniny fermentacyjnej i oczyszczenia (kobalaminy produkuje się tak jak antybiotyki, hodując wyselekcjonowane mikroorganizmy zdolne do ich produkcji), bo jest trwalsza od innych pochodnych (może być przechowywana w formie roztworów w temperaturze pokojowej) i nie reaguje ze składnikami leków i preparatów żywieniowych. A wchłania się równie łatwo jak inne formy i jest w organizmie szybko metabolizowana do pochodnych aktywnych biologicznie.

No i mamy pierwszą aminę! 5’-Dezoksyadenozylokobalamina zalicza się do amin. Ale pozostałe kobalaminy – nie.
W takim razie, nie przedłużając, szybki przegląd pozostałych witamin.
Witamina A
Witaminy z grupy A (retinoidy) to pochodne związków z klasy terpenów – produktów polimeryzacji izoprenu.
Najważniejsze retinoidy to all-trans-retinol, 11-cis-retinal i kwas 9-cis-retinowy. Ponadto w tkankach występują liczne izomery geometryczne, produkty izomeryzacji cis–trans, na przykład all-trans-retinal. Witamerami witaminy A są także karotenoidy, stanowiące prowitaminy A (np.: β-karoten).Izopren

Jak widać, amin w tej grupie witamin nie ma.
Witamina E
Witamina E to nazwa dla grupy aktywnych biologicznie chromanonów, z których najważniejszy jest α-tokoferol. Pozostałe tokoferole (β, γ, δ) różnią się liczbą i położeniem grup metylowych w pierścieniu aromatycznym. Każdy z tokoferoli może występować w postaci ośmiu izomerów, różniących się ułożeniem przestrzennym podstawników wokół trzech chiralnych atomów węgla. Obok tokoferoli znaczenie biologiczne mają nienasycone tokatrienole (każdy z tokatrienoli może występować w postaci dwóch enancjomerów).

Żadna z witamin E nie jest aminą.
Witamina K
Związki z tej grupy to 1,4-naftochinony. Najprostszy z nich, 2-metylo-1,4-naftochinon nosi nazwę witaminy K3. Pozostałe witamery to pochodne z węglowodorowym podstawnikiem w pozycji 3. W roślinach syntezowana jest witamina K1 (filochinon), witamina K2 jest produkowana przez mikroorganizmy, głównie bakterie jelitowe (dominuje zatem w pokarmach odzwierzęcych i produktach fermentowanych).
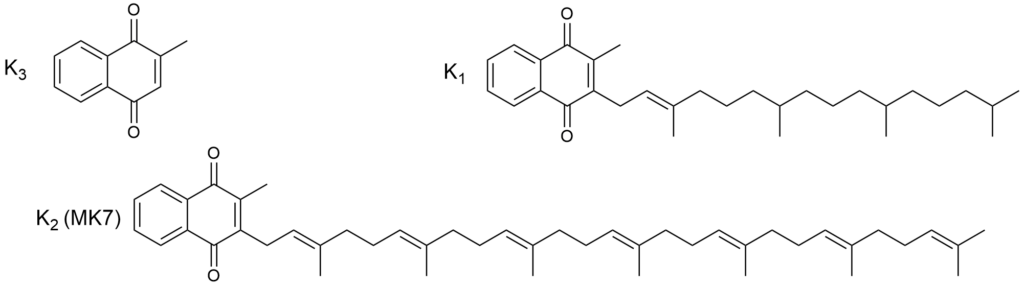
W tej klasie witamin również nie ma amin.
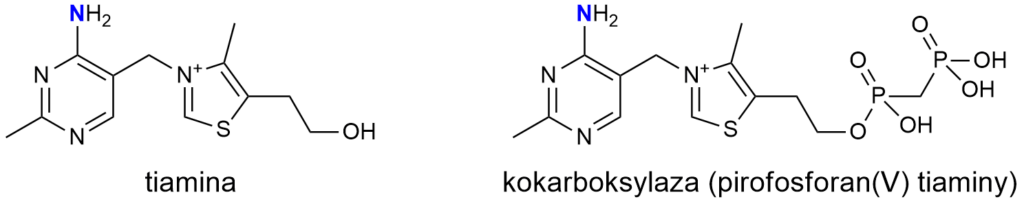
Witamina B1
Witamina B1 (tiamina) to związek otrzymany przez Funka, wykazujący działanie przeciwko chorobie beri-beri. I tak, jest aminą! Grupa aminowa znajduje się w zasadniczej części cząsteczki, tak więc wszystkie witamery z tej grupy są aminami.
Witamina B2
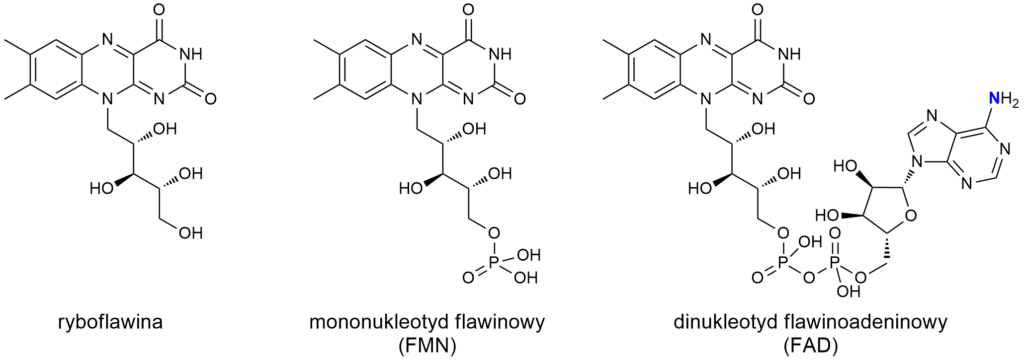

Trzy główne formy witaminy B2 to ryboflawina i jej dwie aktywne pochodne: FMN i FAD. FAD jest aminą, z tym że grupa aminowa pochodzi z adenozyny przyłączonej do fragmentu flawinowego. Ale nie podlega dyskusji, że jeden z witamerów B2 to amina, natomiast ryboflawina i FMN nie są aminami.
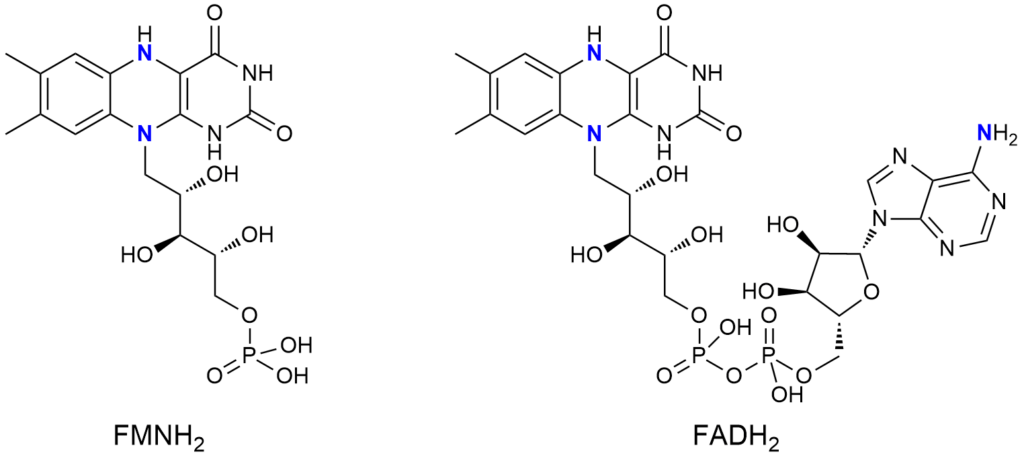
Ale, ale – to nie wszystko. Reszta ryboflawinowa podczas pracy w komórce ulega redukcji. Proces ten zachodzi w obu kofaktorach: FMN i FAD. A taka zredukowana forma (FMNH2 lub FADH2) zyskuje dwa kolejne aminowe atomy azotu.
Witamina B3
Najprostsze formy witaminy B3, czyli kwas nikotynowy i nikotynamid (niacyna) aminami nie są. Natomiast, podobnie jak w wypadku ryboflawiny, przekształcenie w formę aktywną (będącą koenzymem) wiąże się z przyłączeniem do adenozyny. Ponieważ adenozyna jest aminą, to oba koenzymy wywodzące się z witaminy B3 są aminami.

I mamy jeszcze kolejne podobieństwo do kofaktorów flawinowych – redukcja NADP+ lub NAD+ „podczas pracy” prowadzi do wygenerowania aminy.
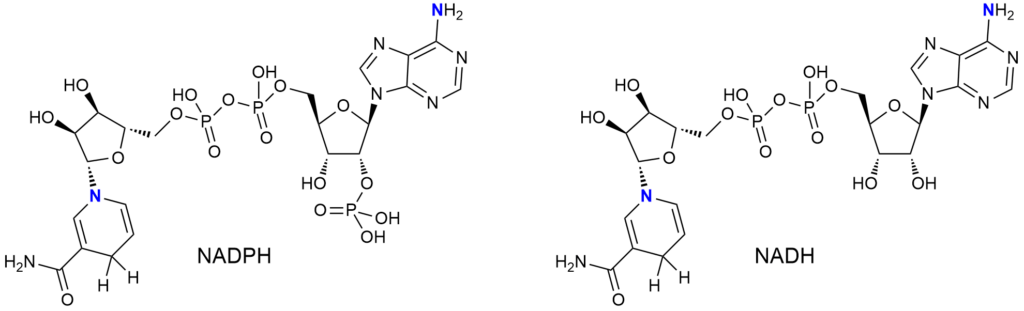
Witamina B5
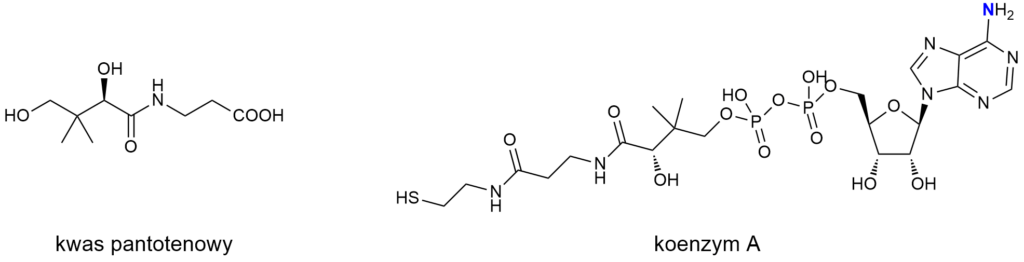
Tak jak w wypadku witamin B2 i B3, jeśli spojrzeć na cząsteczkę witaminy B5 w ścisłym ujęciu, to aminą zdecydowanie nie jest. Cząsteczka kwasu pantotenowego, głównego witameru B5, jest amidem. Natomiast forma aktywna tej witaminy, koenzym A, powstaje przez sprzęgnięcie kwasu pantotenowego z adenozyną, której cząsteczkę zaliczamy do amin. Można więc stwierdzić, że aktywna forma witaminy B5 jest aminą.
Witamina B6
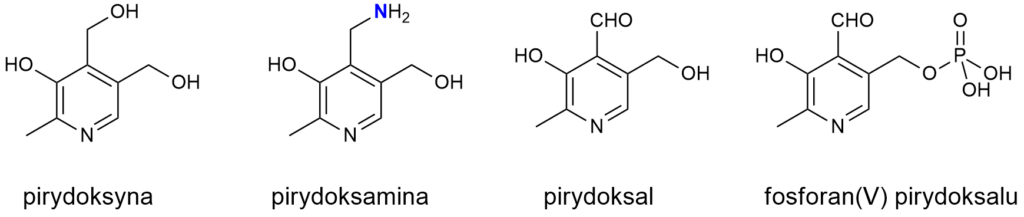
Witamery B6 to pirydoksal, pirydoksyna i pirydoksamina oraz ich estry z kwasem fosforowym(V) (jego cząsteczka przyłącza się do grupy hydroksymetylowej w pozycji 5). Dominującą formą w organizmie jest fosforan(V) pirydoksalu, związany jako kofaktor w centrum aktywnym wielu enzymów. Natomiast pirydoksamina i jej fosforan są aminami.
Witamina B7
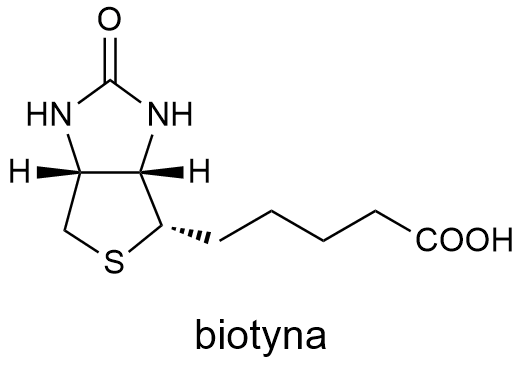
Cząsteczka witaminy B7, biotyna, z chemicznego punktu widzenia jest kwasem karboksylowym, tioeterem i mocznikiem. Aminy się w niej nie doszukamy.
Witamina B9
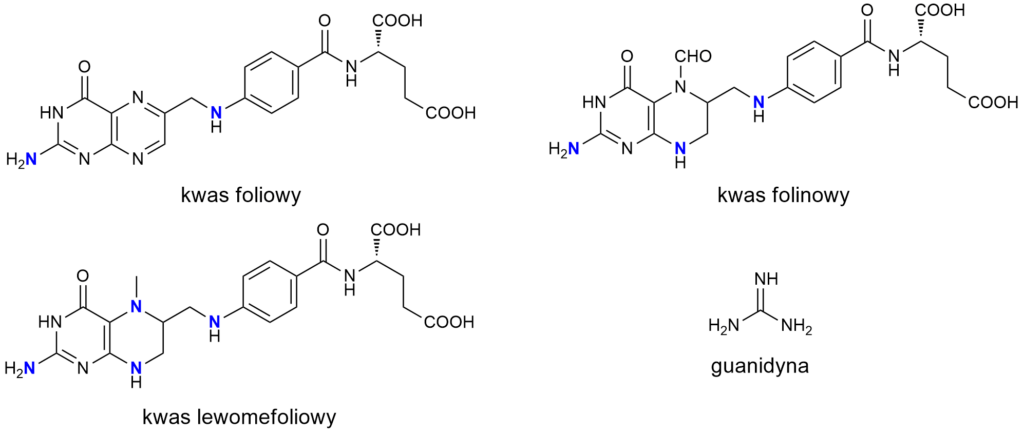
Wszystkie witamery B9 to aminy, przy czym różnią się one liczbą grup aminowych w cząsteczce. Kwas foliowy ma dwie grupy aminowe, kwas folinowy trzy, kwas lewomefoliowy – cztery. Należy podkreślić, że pierwszorzędowa grupa aminowa (-NH2) we wszystkich cząsteczkach stanowi element motywu guanidynowego – więc uznanie jej za aminę może budzić sprzeciw niektórych purystów nomenklatury chemicznej.
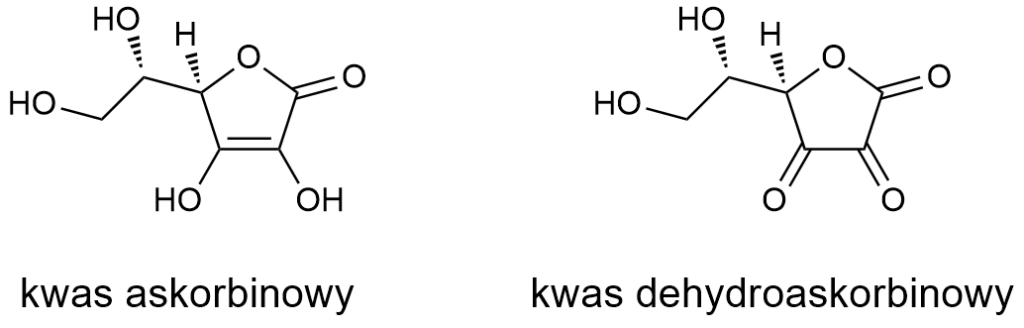
Witamina C
Witamerami w przypadku tej witaminy są kwasy: askorbinowy i dehydroaskorbinowy (oraz ich sole). Wystarczy rzut oka na struktury, aby się przekonać, że aminami nie są.
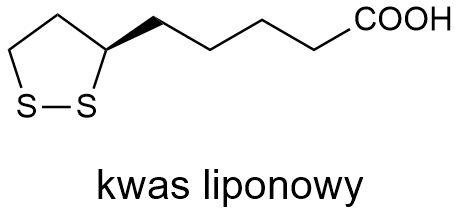
Kwas liponowy
Zacznijmy od tego, że status kwasu liponowego jako witaminy jest dyskutowany. Na pewno jest niezbędny do życia (jako kofaktor enzymów – dehydrogenaz). Ale organizm zwierzęcy, w tym organizm człowieka, radzi sobie świetnie z jego syntezą, zatem nie musi być dostarczany z pożywieniem. Z drugiej strony, witaminę D też sobie syntezujemy z 7-dehydrocholesterolu, który z kolei powstaje z acetylokoenzymu A. Tak czy inaczej, kwas liponowy rodziny aminowych witamin nam nie powiększy.
Dla leniwych
Tym, którym nie chciało się czytać całego tekstu oferujemy esencję w postaci tabeli:

Do poczytania dla zainteresowanych:
(i) P. Moszyński, R. Pyć; Biochemia witamin, Vol. 1 & 2. PWN, Warszawa, 1998.
(ii) A.S. Prasad, G.J. Brewer (eds.); Essential and toxic trace elements and vitamins in human health. Elsevier, London, 2020.
(iii) J. Zempleni, J.W. Suttie, J.F. Gregory III, P.J. Stover (eds.); Handbook of vitamins, 5th ed. Taylor & Francis Group, Boca Raton, 2014.
(iv) G.F.M. Ball; Vitamins: their role in the human body. Blackwell Publishing, Oxford, 2004.
(v) V.R. Preddy (ed.); B vitamins and folate: chemistry, analysis, fFunction and effects. RSC Publishing, Cambridge, 2013.
(vi) K.J. Carpenter; Beriberi, white rice, and vitamin B. A disease, a cause, and a cure. University of California Press, Berkeley, 2000.
(vii) J.I. Young, S. Zuchner, G. Wang; Regulation of the epigenome by vitamin C. Annu. Rev. Nutr., 35: 9.1-9.20, 2015. DOI: 10.1146/annurev-nutr-071714-034228
(viii) C. Funk; On the chemical nature of the substance which cures polyneuritis in birds induced by a diet of polished rice. J Physiol., 43(5): 395-400, 1911. DOI: 10.1113/jphysiol.1911.sp001481
(ix) C. Funk; Beri-beri, polyneuritis in birds, epidemic, dropsy, scurvy, experimental scurvy in animals, infantile scurvy, ship beri-beri, pellagra. J. State Med., 20(6): 341-368, 1912.
(x) G. Jones; Vitamin D analogs. Endocrinol. Metab. Clin. N Am., 39: 447-472, 2010. DOI: 10.1016/j.ecl.2010.02.003
(xi) R.R. Williams; Structure of the vitamin B1. J. Am. Chem. Soc., 58(6): 1063-1064, 1936. DOI: 10.1021/ja01297a515
Komentarze